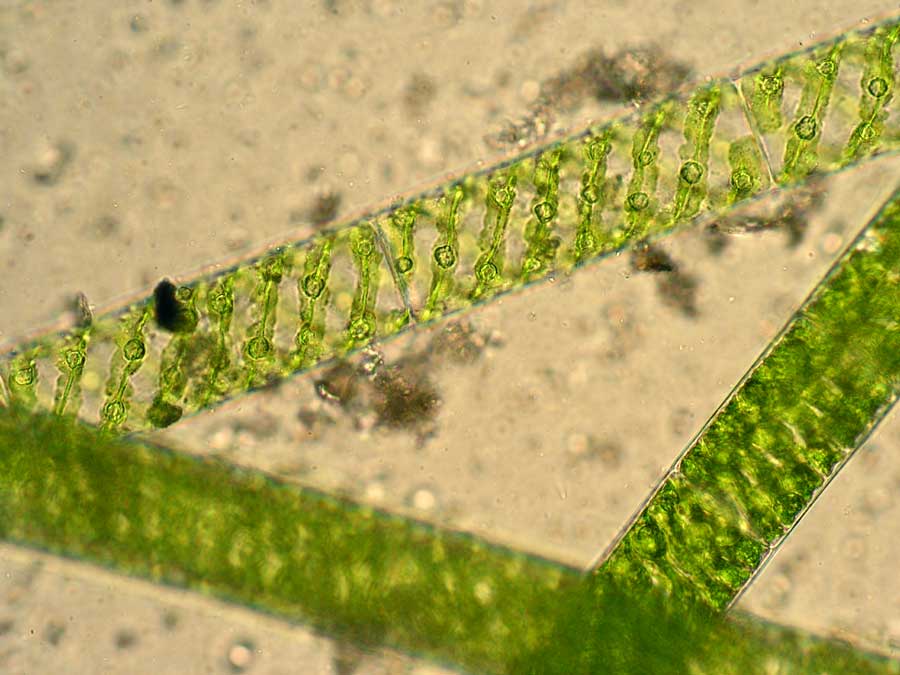
There were other colours of algae in the River Ehen too. Further downstream, several of the stones had a reddish-pink hue which turned out to be composed of a completely different alga. This was Audouinella hermanii, one of the relatively few freshwater species of red algae, more commonly encountered as seaweeds. Like the green algae described in the previous post, this one is also composed of filaments although this time they have a red- rather than green colour. There are some short side-branches bearing clusters of egg-shaped “carpogonia” (the female reproductive organs) and others which bear, like Draparnaldia, long, colourless hairs. These, too, probably have a role in the phosphorus nutrition of the alga.


The left hand image shows a cobble covered with Audouinella from the River Ehen, whilst the right hand image shows filaments and carpogonia under the microscope. The filaments are approximately 10 micrometres (a hundredth of a millimetre) in diameter.
Some of the filaments also had short filaments of a blue-green alga, called Heteroleibleinia (formerly Lyngbya) rigidula growing on them and there were also many diatoms, particularly a species of Gomphonema growing on long stalks in and around the Audouinella filaments. Later in the year, these growths of Audouinella get so completely smothered by diatoms that they lose their reddish appearance and become dark brown.


The left hand image shows filaments of Heteroleibleinia rigidula growing on the Audouinella filaments; the right hand image shows four cells of the Gomphonema species (three in valve view and one, on the right, in girdle view).
The final image is a diorama putting all of these components together, with four distinct groups of algae in close juxtaposition. The Audouinella filaments dominate the view but there are three diatoms, on their long stalks on the left hand side, along with a few filaments of Heteroleibleinia rigidula. In the bottom right corner there are a few cells of Stigeoclonium tenue, a relative of Draparnaldia that I also saw growing amidst the Audouinella filaments. Several of the Audouinella filaments end in colourless hairs on the right hand side. The colour of the Audouinella reflects the pigment composition of the cells: the green of the chlorophyll is masked by two other pigments: phycocyanin, which is blue, and phycoerythrin, which is red. A mix of Hooker’s Green, Ultramarine and Crimson Alizarin gives an approximation of the hue. There should be several ribbon-shaped chloroplasts lying just inside the cell wall, but these were hard to resolve with a light microscope.

A three-dimensional representation of the Audouinella-dominated community in the River Ehen, February 2013.
Brian Whitton identified the Heteroleibleinia rigidula for me.